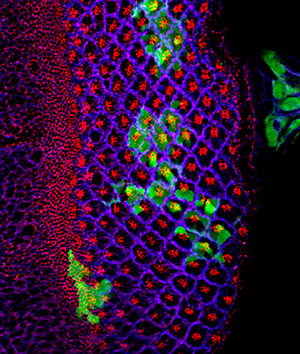
Genetics of Development and Regeneration
Florenci SerrasUnderstanding how cells coordinate proliferation, differentiation, and organization to form functional tissues is a central challenge in developmental biology. Remarkably, similar processes are reactivated during tissue regeneration, where cells must rebuild complex structures after damage. A key question in modern biology is how the genetic programs that drive regeneration compare to those that guide normal development—what is shared, and what is unique?
Our research aims to identify the genetic programs that enable tissue regeneration and to uncover the mechanisms through which they operate. To address these questions, we use Drosophila imaginal discs—epithelial tissues that provide a powerful and versatile model system for genetic analysis. By combining precise genetic tools with targeted tissue injury, we can disrupt epithelial integrity and directly observe the processes of repair and regeneration in vivo.
The development of engineered genetic tools in imaginal tissues has been instrumental in revealing fundamental principles of epithelial regeneration. Through this approach, our work seeks to bridge the gap between development and regeneration, providing insights that may ultimately inform regenerative medicine.
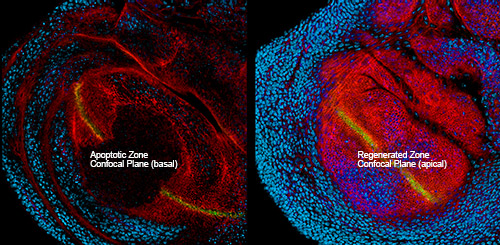
We genetically activate cell death to induce regeneration (Bergantiños et al 2010). The technique is based on the activation of apoptosis in specific zones using tissue specific transactivation drivers controlled by temperature sensitive alleles.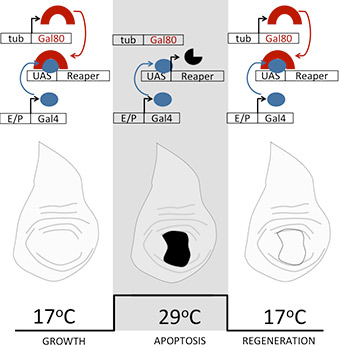
This is an alternative technique that allows two cell populations in the same imaginal disc to be genetically programmed to express different transgenes (Santabárbara-Ruiz et al., 2015). One transactivator can drive the pro-apoptotic gene (e.g. rpr or hid) whereas the other can express a tester transgene (e.g. a UAS-RNAi for a gene of interest). Using this strategy, the combination of lexA and Gal4 double transactivator system has been key for our regeneration studies. LexA is a bacterial transcription factor that binds to specific sequences known as the LexA operator or lexO. An optimized LexA transactivation system has been engineered to work together with Gal4. Briefly, chimeric proteins have been created in which LexA (L) contains the activation domain of yeast Gal4 (G) separated by a hinge region (H), which results in a Gal80TS suppressible LHG (Yagi, Mayer, and Basler 2010).
Activated LHG induces the expression of any transgene under the control of the lexO sequences and does not interfere with Gal4/UAS. For combined transactivation systems, the LHG/lexO-rpr is used to induce cell death in specific domains of the imaginal disc, and the Gal4/UAS system to activate the desired UAS-transgene (UAS-RNAi, UAS-mutated forms, etc), both controlled by the temperature sensitive allele Gal80TS. This combined system has allowed to monitor gene function in regeneration. We aim to improve these genetic designs for ex vivo imaging.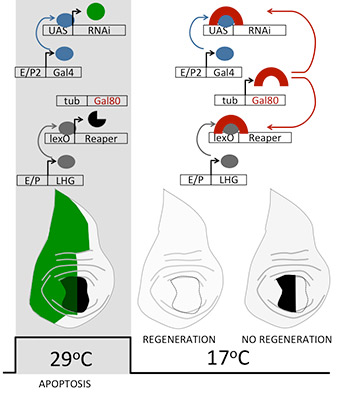
Another method is to inflict a physical injury in discs and analyze the response to damage after implantation in the female abdomen, which acts as a culture chamber.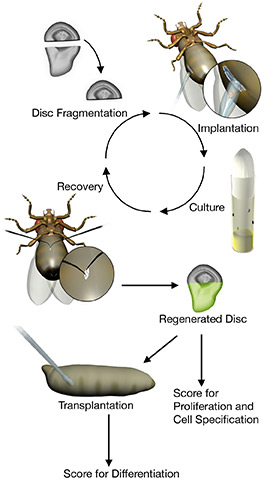
Alternatively, discs can be cultured ex vivo after physical injury and imaged. The use of fluorescent reporters helps to observe gene activity in living tissues.
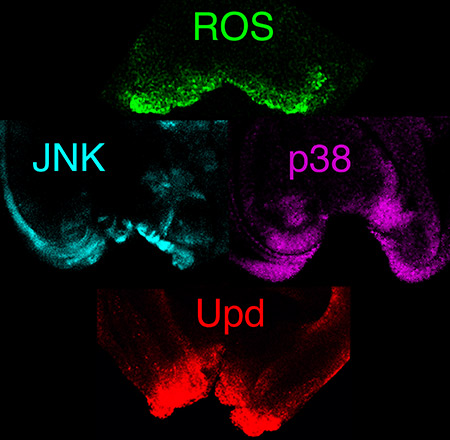
An important issue in regeneration research is to discover the early signals that trigger regeneration. Physical damage or induction of cell death result in a burst of reactive oxygen species (ROS) that spreads to neighboring healthy tissue. This event induces activation of the p38 and JNK stress activated protein kinases.
Downstream of these MAPKs, the leptin-like cytokines Unpaired (homologous to the human IL-6) are transcriptionally activated. These bind to the IL-6R type receptor that activates a Janus kinase, and thereby promotes the translocation of a STAT3-like transcription factor STAT92E to the nucleus.
We are currently analyzing how these stress responses are sensed into the cells near a damaged zone in order to be recruited to start healing and regeneration.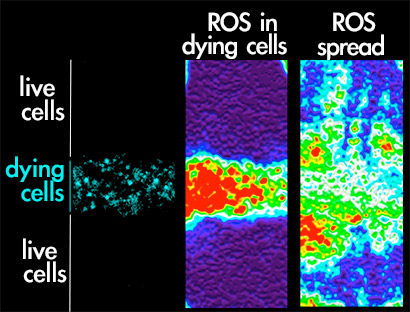
To regenerate a missing structure, epithelial cells must be able to re-specify their fates to reconstruct the missing tissue. Thus, our goal is to understand how cells can switch from one genetic program to another. To this aim, we investigate the genetic and epigenetic basis of cell fate re-specification during regeneration.

During normal growth, cells divisions are oriented to coordinate tissue size and shape. In addition to proliferation and re-specification, epithelial regenerating cells replace the missing tissue by activating proliferation and reorienting cell divisions to repopulate the missing zones.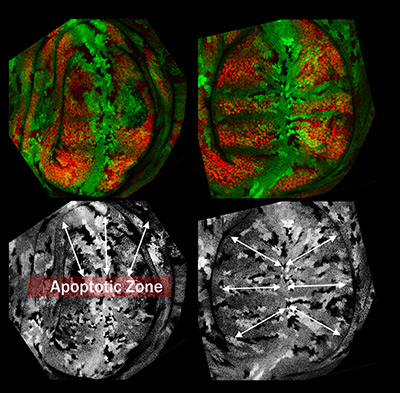
A single cell resolution model to examine the relationship between cell signaling and cell-fate specification is the R7 photoreceptor in the Drosophila retina. The activation of the Sevenless receptor tyrosine kinase, which upon activation signals through Ras/MAPK to the nucleous, plays a pivotal role in the specification of the R7 cell. Although that pathway has been well characterized, its regulators and links to other signaling pathways are poorly known. Genetic screenings and transcriptome analysis have been used in the lab to discover the genes that respond to different levels of signaling activity.